|
|
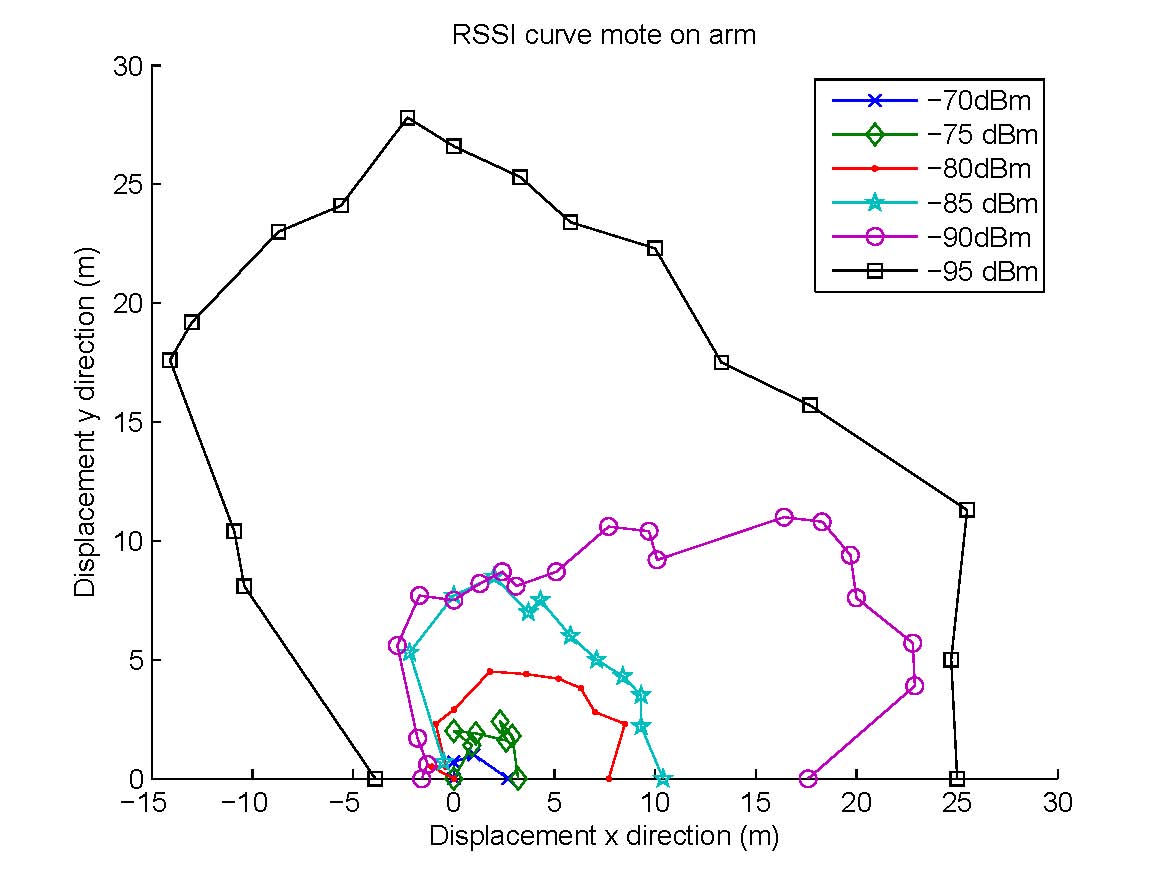
In choosing the preferred mounting position several aspects have to be considered: the attenuation of the wireless signal by the body, the ease and stability of attachment, and the possibility of damage to the device itself. We conducted experiments to profile the propagation of wireless signals around the body in an open field, and show in the Figure above a rough map of the received signal strength indicator (RSSI) at various distances and directions from the body-mounted mote device, which transmits data every second at the highest available power level of 1mW. The person wearing the device is located at (0,0) and is facing north. The Figure shows the RSSI contours when the device is attached to the right arm. Not surprisingly, the reach is much larger to the right (at around 16m the signal fades to below -95dBm) than to the left (signals barely reach beyond 3m), and there is a 5-6m reach to the front and back of the player. By contrast, when the device is mounted on the back, we found that the contours stretch up to 16m to the back, but barely a couple of metres to the front. The smaller wireless ``shadow'' for the arm attachment can be attributed to the smaller cross-section of the body presented to the signal wave. Other reasons also favour an arm-mounted position, such as absence of clothing impediments, and less chance of injury/damage during a fall. These considerations led us to use an arm-mounted position for all our field studies. |
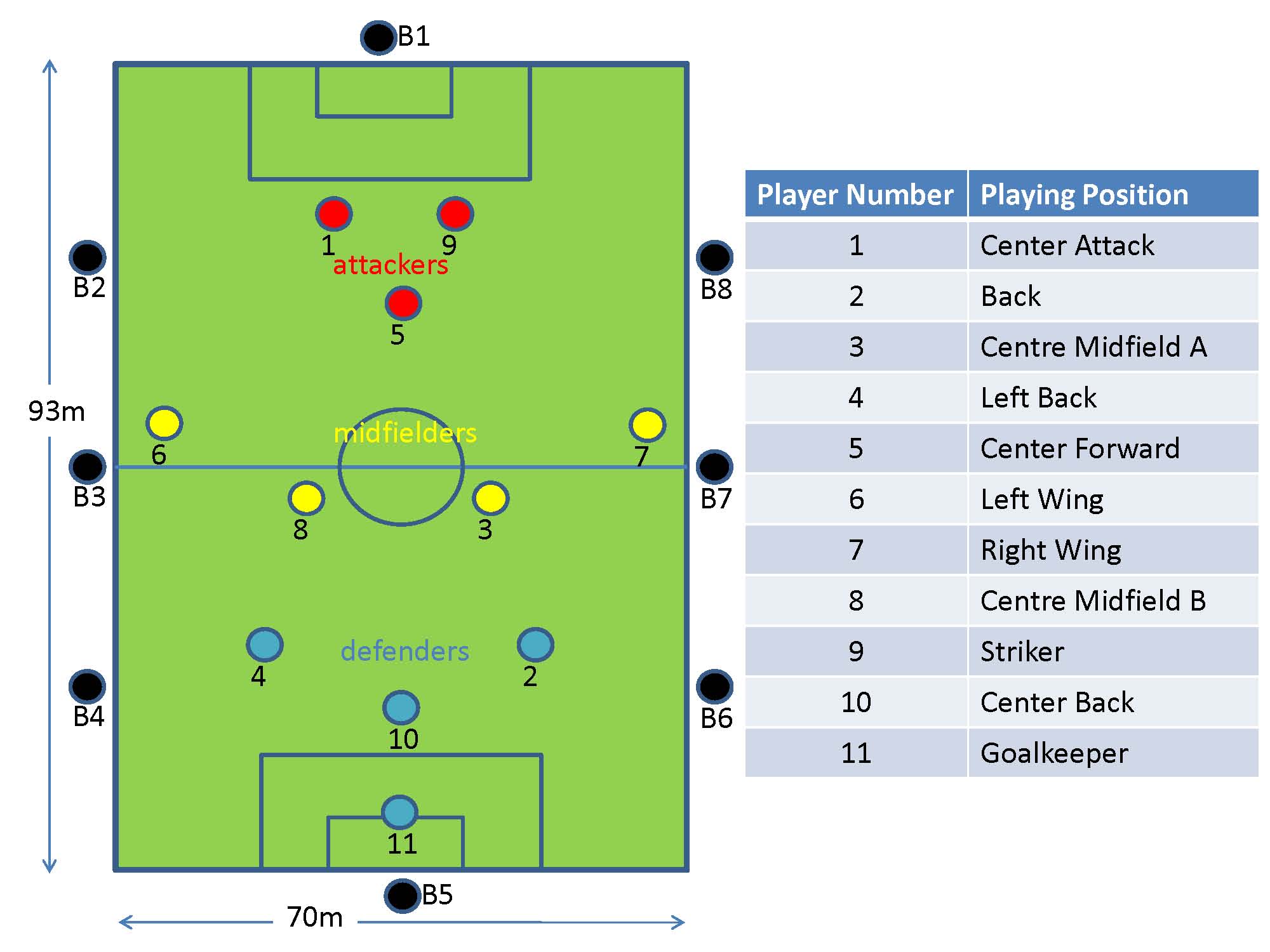 |
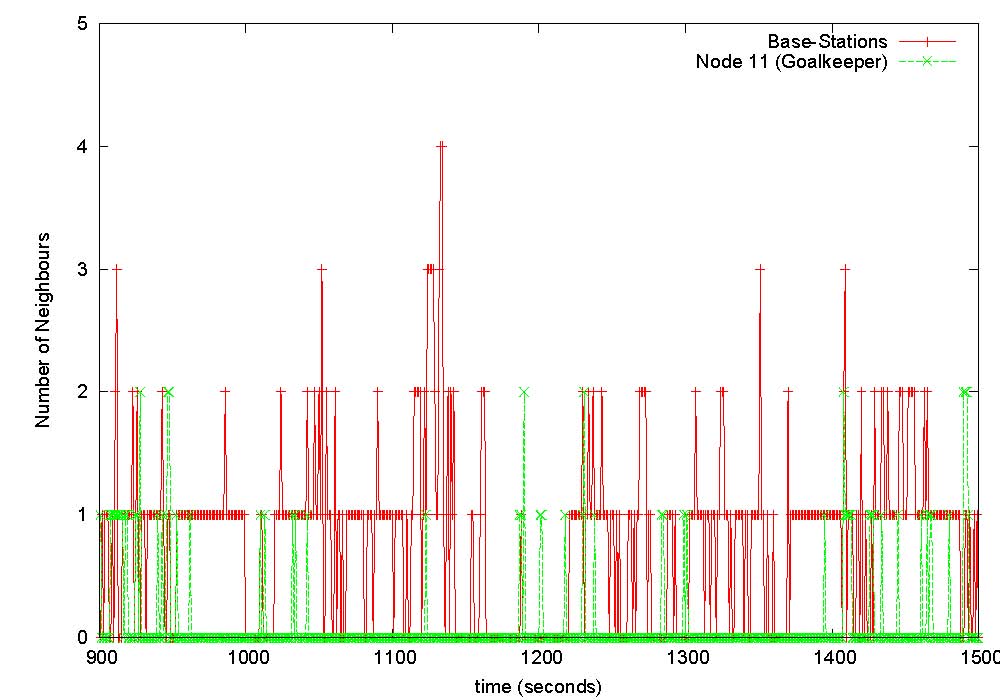
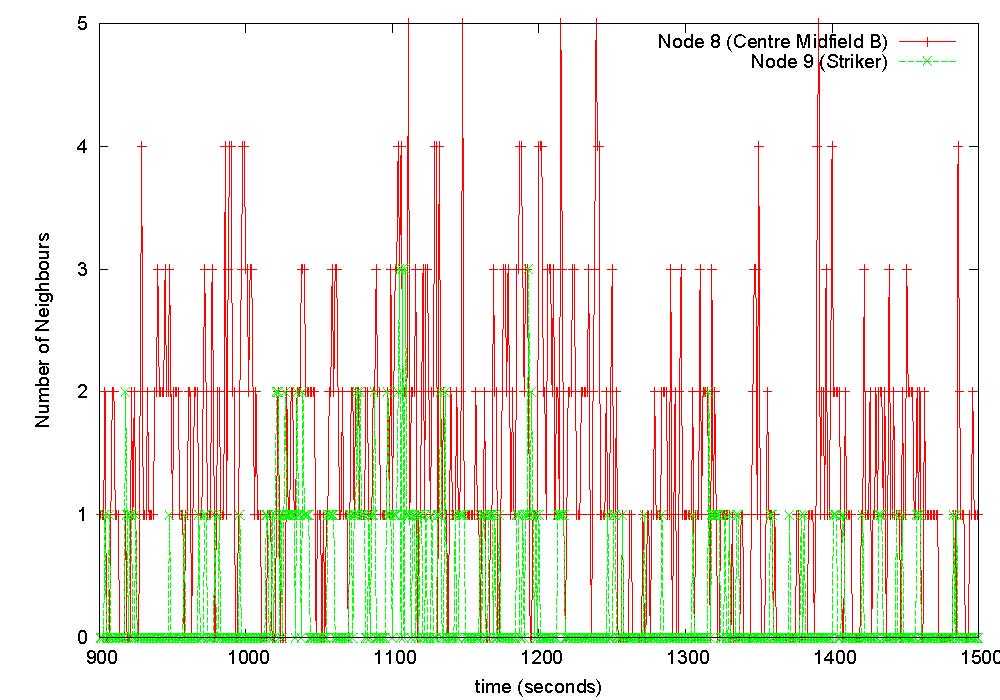
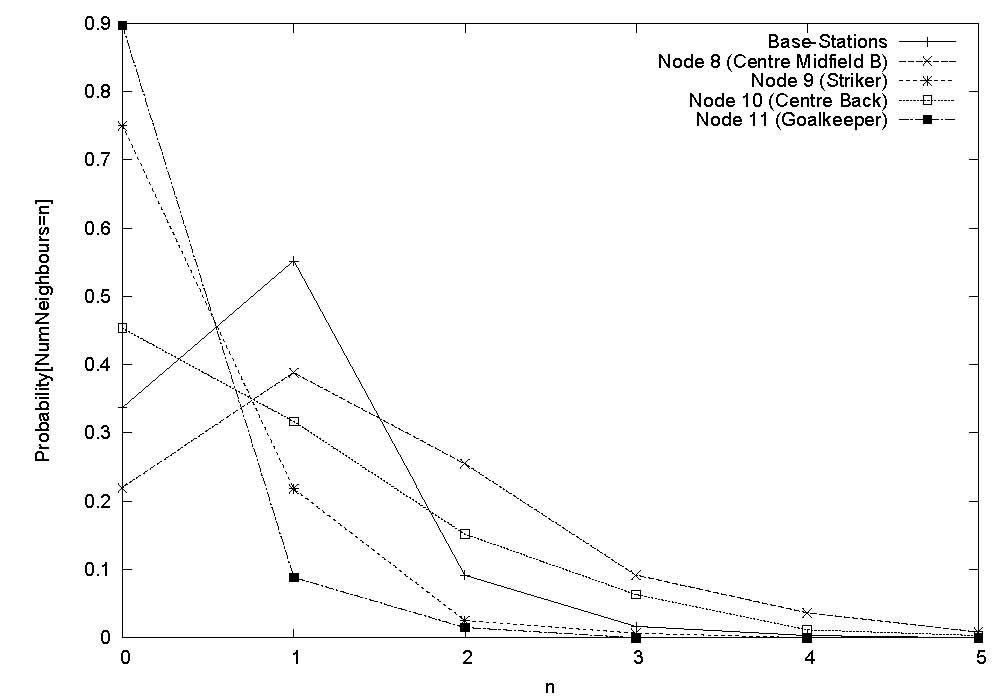
The game was played on a full size field with dimensions 93m x 70m. Each of the 11 players wore a monitoring device on their arm, and 8 base-stations were positioned (at a height of about 1m from the ground) along the sidelines of the playing area. The figure shows the nominal playing positions and associated node identification numbers. Unfortunately the devices worn by players 2 (back) and 4 (left back) were damaged during play and we could not obtain data from them, as was base-station B3 which got hit by the ball. We implemented software on each of the body-worn devices such that it broadcasts, once every second, at the highest available power level of 1mW, a packet containing its unique identifier and a sequence number, during the entire measurement period. All devices (body-worn as well as base-stations) that successfully receive this packet record this event in their on-board memory. As the game proceeds, each node (and base-station) will be cataloguing which other nodes it could hear at each time instant. To prevent collisions in-the-air, each second is divided into 11 slots each of approximate duration 90ms, and each of the 11 body-worn devices is given a unique such slot for transmission every second. Just prior to commencement of the game, the master base-station sends a clock synchronisation message to all nodes, upon receipt of which each node starts recording connectivity data in on-board memory. Data collection stops after 25 minutes, and at the end of the game data from each node is extracted by the master base-station for off-line analysis. |
 |
 |
 |
|
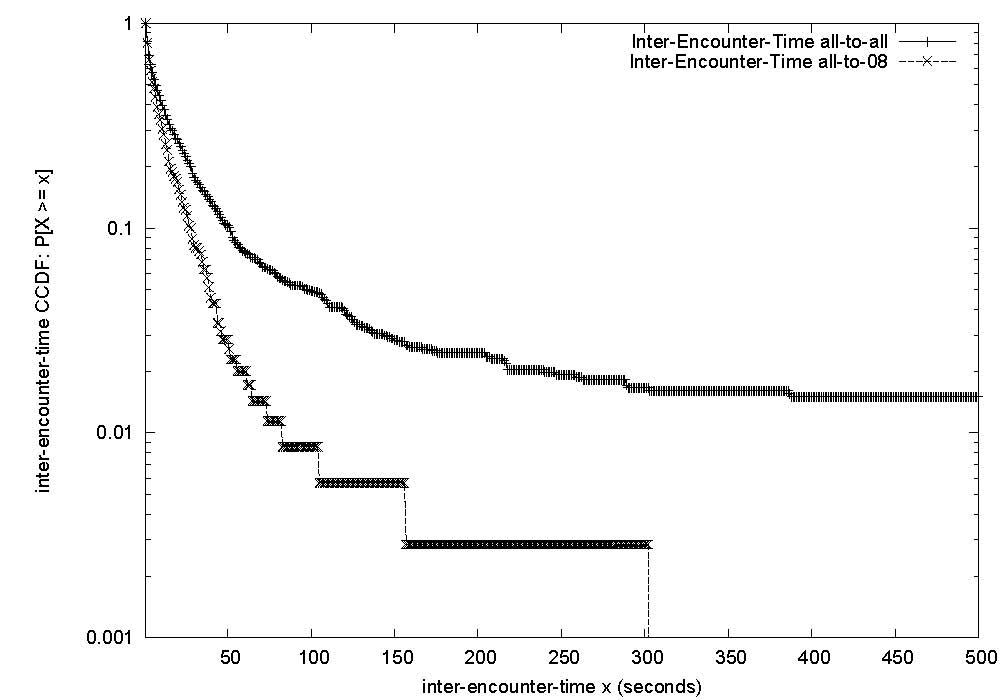 |
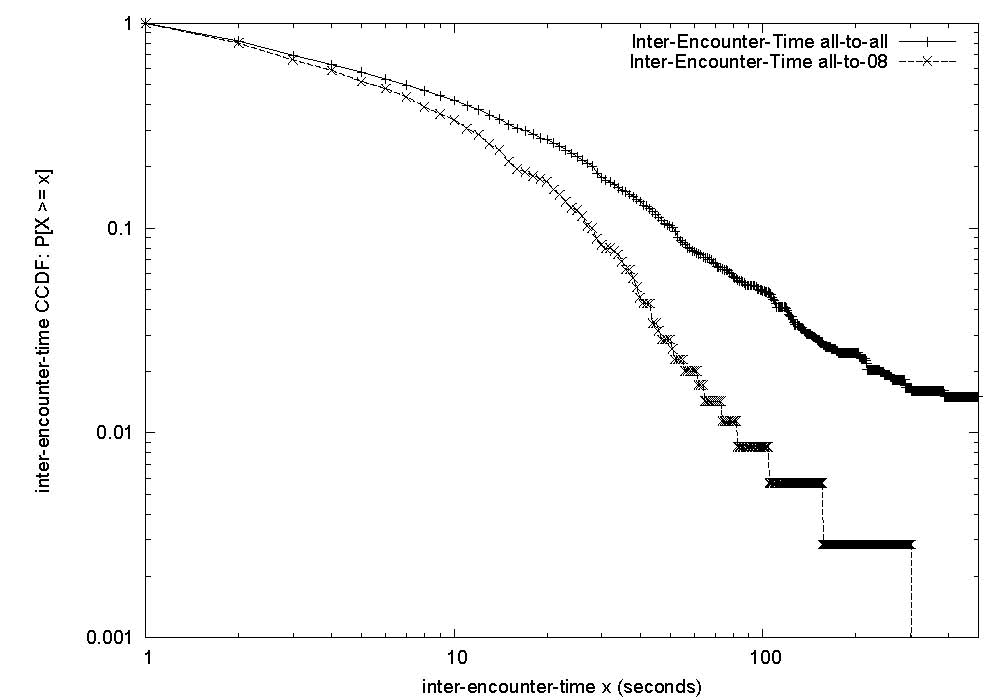 |
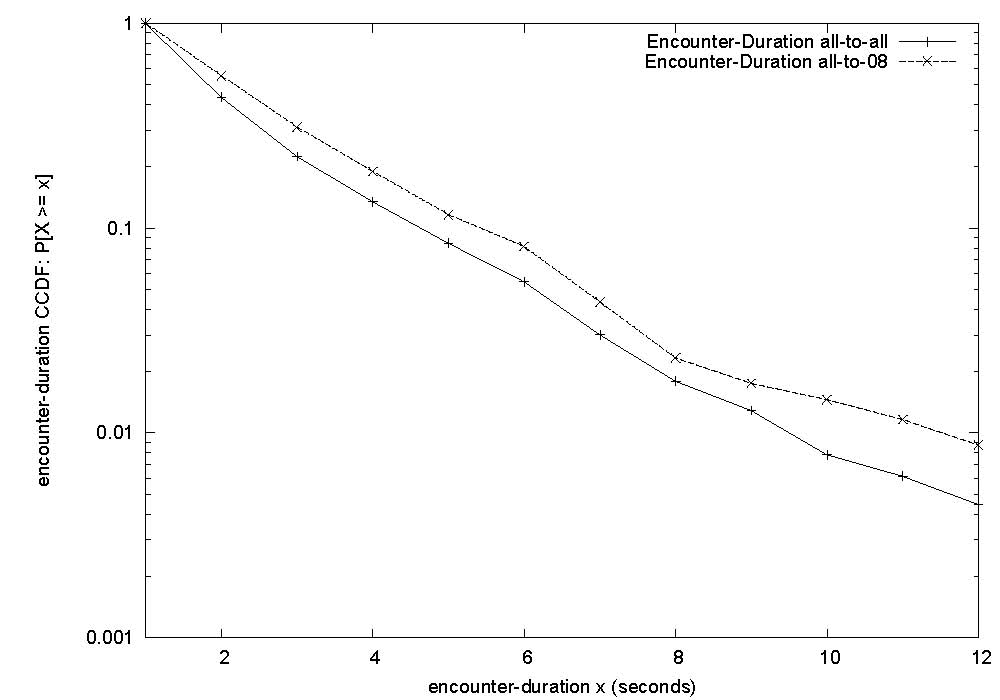 |
 |
 |
 |
 |
 |